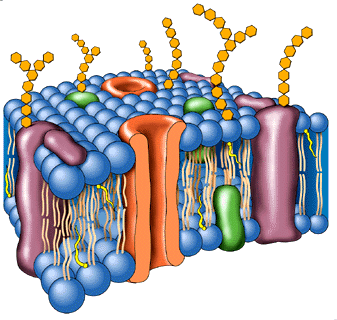
Receptores transmembrana
Los receptores transmembrana son proteínas que se extienden por todo el espesor de la membrana plasmática de la célula, con un extremo del receptor fuera de la célula (dominio extracelular) y otro extremo del receptor dentro (dominio intracelular). Cuando el dominio extracelular reconoce a una hormona, la totalidad del receptor sufre un cambio en su conformación estructural que afecta al dominio intracelular, confiriéndole una nueva acción. En este caso, la hormona (u otro ligando) no atraviesa la membrana plasmática para penetrar en la célula. Aunque un receptor sencillo puede transducir alguna señal tras la unión del ligando, lo más frecuente es que la unión del ligando provoque la asociación de varias moléculas receptoras. Los principales tipos de receptores transmembrana son los siguientes:1
[editar]Receptores con actividad tirosina quinasa intrínseca
Dentro de este grupo están los receptores de la mayor parte de los factores de crecimiento, como EGF, TGF-alfa, HGF, PDGF, VEGF, FGF, y el receptor de la insulina. Los receptores de esta familia tienen un dominio extracelular de unión al ligando, un dominio transmembrana, y un dominio intracelular con actividad tirosina quinasa intrínseca. Cuando se une el ligando, el receptor se dimeriza, lo que induce la autofosforilación de las tirosinas del dominio intracelular y activa la tirosina quinasa, que fosforila (y por tanto activa) muchas moléculas efectoras en cascada, de forma directa o mediante proteínas adaptadoras. Estos receptores pueden activar cascadas de señalización diferentes, como por ejemplo:
- la cascada de las MAP quinasas (por mitogen-activated protein), con activación de la proteína de unión a GTP denominada Ras, y síntesis y activación de factores de transcripcióncomo FOS y JUN, que estimulan la producción de nuevos factores de crecimiento, de receptores para dichos factores y de proteínas que controlan la entrada de la célula en el ciclo celular
- la cascada de la PI3K (fosfoinositol 3-quinasa), que activa la quinasa Akt, implicada en proliferación celular y supervivencia celular por inhibición de apoptosis
En muchos tipos de cáncer se han detectado alteraciones en la actividad tirosina quinasa del receptor y mutaciones, por lo que estas moléculas son dianas terapéuticas muy importantes.
[editar]Receptores que carecen de actividad intrínseca y reclutan quinasas
En este grupo se incluyen los receptores de muchas citoquinas, como IL-2, IL-3, interferón α, β y γ, eritropoyetina (EPO), hormona del crecimiento y prolactina. La transmisión de la señal de estos receptores provoca la activación de miembros de la familia de quinasas denominadas JAK (Janus quinasas). Estas quinasas activan factores de transcripción citoplásmicos llamados STATs (por signal transducers and activation of transcription), que se translocan al núcleo y activan la transcripción de genes específicos. En otros casos, estos receptores activan la cascada de las MAP-quinasas.
[editar]Receptores asociados a proteínas G
En este caso, la transducción de la señal se realiza a través de proteínas triméricas de unión a GTP (proteínas G), que constan de 7 hélices transmembrana y constituyen la mayor familia de proteínas receptoras (1% del genoma humano). Hay un gran número de ligandos que utilizan estos receptores, como las quimiokinas, vasopresina, serotonina, histamina,adrenalina, noradrenalina, calcitonina, glucagón y hormona paratiroidea, entre otros. Muchas drogas farmacéuticas comunes tienen como diana estos receptores. La unión del ligando provoca cambio de conformación y activación del receptor, que puede interaccionar con otras muchas proteínas G. La forma inactiva une GDP, mientras que la forma activa une GTP. En algunos casos, esta vía de señalización incluye AMPc como segundo mensajero.
Segundos Mensajeros
Los efectos de la unión de muchas de estas hormonas están mediadas en el interior celular por la regulación en la concentración de otras moléculas de bajo peso molecular denominadas segundos mensajeros. Estas moléculas incluyen al AMPc, GMPc, diacilglicerol (DAG), 1,4,5-inositol trifosfato (IP3), varios fosfolípidos de inositol y el calcio (Ca+2).
La concentración de estas moléculas incide en la regulación del metabolismo celular, actividad enzimática o no enzimática de proteínas, y transcripción de genes específicos implicados en la proliferación, diferenciación y supervivencia celular, además de proporcionar a la molécula señalizadora una forma de transducción y amplificación de esta señal en el interior celular. En este aspecto de la señalización profundizaremos en adelante.

bibliografia: http://es.wikipedia.org/wiki/Receptor_celular





 (e.g. porinas, proteínas con estructura en forma de barril construido de 8 a 22 láminas
(e.g. porinas, proteínas con estructura en forma de barril construido de 8 a 22 láminas